Ģenētiskā dreifēšana: kas tas ir un kā tas ietekmē bioloģisko evolūciju?
Bioloģiskā evolūcija, kas iecerēta kā iedzimtu īpašību izmaiņu kopums 2005 paaudžu paaudzē dzīvojošās būtnes ir pats dzīves un pielāgošanās jaunajam dzinējs videi.
Dzīvo būtņu populācijas variāciju nosaka virkne DNS mutāciju, kas notiek nejauši, tas ir, tas nereaģē uz apzinātu mehānismu. Pat ja tā, šo mutāciju atlases procesi var būt nejauši, vai gluži pretēji, tiem ir pilnībā pamatots izskaidrojums.
Tādējādi evolūcija ir spēks, kuru uztur nejaušas mutācijas, ģenētiskā rekombinācija laikā seksuālā reprodukcija un gēnu plūsma (jaunu dalībnieku ienākšana populācijā), starp daudziem citiem faktoriem. Viens no šiem būtiskās nozīmes faktoriem, kas bieži vien izvairās no vispārējas izpratnes, ir termins, kas mūs šeit skar: ģenētiskais dreifs. Šeit mēs izskaidrojam visu par šo aizraujošo procesu.
- Saistītais raksts: "Bioloģiskās evolūcijas teorija"
Kas ir ģenētiskais dreifs?
Pirmkārt, mēs varam definēt šo sarežģīto terminu kā "evolucionāru spēku, kas darbojas kopā ar dabisko atlasi, laika gaitā mainot sugu alēļu frekvences". Kā priekšvārds jāatzīmē, ka
tas ir stohastisks process, tas ir, tas notiek nejaušības vai sporādisku nekorelētu efektu dēļ.Papildus šim sākotnējam raksturlielumam vēl viens termins, kas nosaka ģenētisko novirzi, ir zaudējums, jo tas ir selekcijas spēks veicina alēļu variāciju populācijā, dažus fiksējot un veicinot izzušanu citi. Mēs to sīkāk redzēsim nākamajās rindās.
Par alēlēm un populāciju
Ģenētisko terminu vienkāršošana, mēs varam apstiprināt, ka alēle ir katra no variācijām, kuras var uzrādīt viens un tas pats gēns. Klasisks piemērs tam ir zirņu sēklas, kuras Mendels izmantoja, lai izskaidrotu ģenētisko segregāciju paaudzēs. Dominējošā "A" alēle var kodēt zaļo sēklu krāsu, bet recesīvā "a" alēle - dzelteno krāsu.
Tā kā lielākā daļa dzīvnieku ir diploīdas būtnes (kuru kodolā ir divas homologu hromosomu kopas), katra no šīm divām kodējošajām alēlēm raksturs nāks attiecīgi no tēva un mātes, tāpēc iespējamās variācijas šajā gadījumā būtu šādas: AA, Aa un aa. Tādējādi, ja mēs saprotam, ka indivīds katram gēnam pārmanto divas alēles, nāks viņa fenotips (ārējās īpašības) ko tieši kodē viņu genotips (alēļu kombinācijas viņu genomā), kas tiek pārmantots kā Viņa vecāki.
Otrkārt, termins “populācija” ir nedaudz jāpēta bioloģijas jomā, tā kā ģenētiskā novirze iedarbojas uz populācijām, nevis uz pašu sugu. Suga ir "slēgts" jēdziens, jo tā nevar apmainīties ar gēniem ar citām atšķirīgām vienībām. No otras puses, iedzīvotāji tiek uzskatīti par “atvērtiem” nodalījumiem, jo tajā var iekļūt dažādi citu locekļu locekļi. populācijām, bet tās pašas sugas un vairojas to vidū, notikumam, kam būs izšķiroša nozīme līnijās vēlāk. Kad mēs vispārīgi esam izveidojuši abus terminus, mēs esam gatavi saprast ģenētiskās dreifēšanas pamatu.
- Jūs varētu interesēt: "Specifikācija: kas tas ir un kā tas attīstās bioloģiskajā evolūcijā"
Dreifa teorētiskais pamats
Ir pienācis laiks turēties pie sēdekļa, jo līknes un termini ir nedaudz sarežģīti izskaidrojami. Ģenētiskais dreifs nosaka alēliskās frekvences dispersija, tas ir, rakstzīmju mainīgums attiecībā pret vidējo. Tādējādi mēs varam aprēķināt šo evolūcijas spēku, izmantojot šādu formulu:
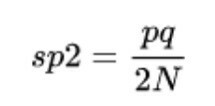
- sp2 atbilst populāciju alēļu frekvenču dispersijai, tas ir, pašam ģenētiskajam dreifam.
- p un q ir rakstura divu populāciju alēļu frekvences.
- N ir indivīdu skaits katrā no divām populācijām.
Protams, katrs no šiem parametriem tiek iegūts, izmantojot sarežģītas formulas, tāpēc mēs nepievērsīsimies vairāk uzmanības šī evolūcijas spēka matemātiskajam pamatam. Ja pēc šo rindiņu izlasīšanas idejai ir jābūt skaidrai, tā ir šāda: jo mazāks ir populācijas lielums, jo lielāka būs ģenētiskā novirze pār tās locekļiem.
Faktiskais populācijas lielums
Iepriekšējā rindkopā mēs esam ieviesuši galveno terminu: populācijas lielums. Patiesība ir tāda, ka, ņemot vērā ģenētiskās novirzes lielumu, zinātniekiem ir vērts skaitīt ne tikai populācijas indivīdus. Šajos gadījumos tajā droši reproducējamo dzīvnieku skaits.
Ļoti skaidrs atšķirības starp kopējo iedzīvotāju skaitu un faktisko populācijas lielumu piemērs ir abinieku demogrāfiskie pētījumi. Piemēram, kopēju krupju populāciju var veidot 120 locekļi. Ja mēs izmantojam ģenētisko analīzi, mēs varam novērot, ka noteikti tikai aptuveni 40 pieaugušie pieaugušie gadā vairojas, atstājot maksimāli pēcnācējus. Tādējādi faktiskais populācijas lielums (Ne), kas cieš no dreifēšanas, būtu 40, nevis 120.
Ģenētiskā dreifa sekas
Ģenētiskajam dreifam ir vairākas ietekmes uz dzīvo būtņu populācijām, taču mēs varam tos sadalīt divos lielos blokos:
- Izmaina alēles biežumu populācijā. Tas var nozīmēt, ka šie rādītāji palielinās vai samazinās, jo tas ir tīra gadījuma jautājums.
- Samazina ilgtermiņa ģenētiskās variācijas populācijās.
Šis pēdējais punkts ir ārkārtīgi svarīgs, jo ģenētiskā novirze samazina mainīgumu, kas galu galā izpaužas kā lielāka iedzīvotāju neaizsargātība pret vides izmaiņām. Ņemsim praktisku piemēru.
Ja mūsu fiktīvajā populācijā ir 10 putni, 8 sarkanie un 2 dzeltenie, ir dabiski domāt, ka tīra gadījuma dēļ tas ir ticamāk nekā nākamās paaudzes laikā sarkanie dalībnieki ir vairāk pārstāvēti (jo, ja no šiem 10 atveido tikai 3, pastāv iespēja, ka visi 3 ir krāsaini Sarkans). Pirmajā paaudzē sarkanās rakstzīmes "p" alēliskā frekvence būtu 0,8, bet dzeltenā "q" - 0,2.
Ja notikumā vairojas tikai 3 sarkanās krāsas tēviņi un mātītes, teorētiski alēle q varētu pazust nākamā paaudze, tātad p = 1 un q = 0, visiem pēcnācējiem ir sarkana krāsa (burts p būtu bijis komplekts). Tas ir ģenētiskā dreifa reālais efekts, kas nejauši tas rada visvairāk izplatīto rakstzīmju fiksāciju populācijā un galu galā atmet unikālākos.
Iedzīvotāju glābšana
Par laimi, mums ir spēks, kas lielā mērā izvairās no šīs izlases izvēles: dabiskā atlase. Šajā gadījumā, mēs saskaramies ar evolūcijas dzinēju, kas nepavisam neatbilst nejaušiem un stohastiskiem procesiem, jo katra indivīda īpašības var noteikt viņu izdzīvošanu, vairošanos un sekojošo pārstāvību nākamajās paaudzēs.
Jāatzīmē arī tas, ka iepriekš minētais piemērs ir diezgan klibs pašnoteikta redukcionisma dēļ, tā kā nepārprotami daudzas morfoloģiskās rakstzīmes kodē vairāk nekā viens gēns (piemēram, acu krāsa) piemērs). Turklāt 1000 indivīdu, nevis 10 cilvēku populācijā ir skaidrs, ka alēles izzušana ir daudz sarežģītāka nekā tās "dzēšana" vienā paaudzē.
No otras puses, gēnu plūsma ir vēl viens galvenais jēdziens, kas ļauj izvairīties no ģenētiskās novirzes. Alēli populācijā laika gaitā varēja noteikt, bet, ja parādās jauni dalībnieki ar dažādām alēlēm un vairoties kopā ar sākotnējās populācijas indivīdiem, atjaunotā ģenētiskā mainība tiek ieviesta turpmāk paaudzēm.
Visbeidzot, tas ir jāierobežo mutācijas notiek nejauši dzīvās būtnēs. Tādējādi DNS, kas kodē jaunas alēles, var rasties variācijas, tāpēc (plkst mazāk teorētiski) slēgtā populācijā jaunas rakstzīmes var turpināt parādīties a sporādiski.
Turpināt
Kā mēs redzējām, ģenētiskā dreifēšana tas ir galvenais dzīvo būtņu evolūcijas dzinējs kopā ar dabisko atlasi, bet tas atšķiras no pēdējiem nejaušības un nejaušības dēļ. No tīri teorētiskā viedokļa, ja nebūtu tādu notikumu kā gēnu plūsma, mutāciju parādīšanās vai dabiskā atlase, visām populācijām katram gēnam būtu viena alēle, pat ja tas prasītu daudz paaudzēm.
Tas, protams, izpaužas kā mazāka ģenētiskā mainība, kas nozīmē sliktāku iedzīvotāju un indivīdu līmeņa reakciju uz izmaiņām un vides neuzmanību. Tādējādi ģenētisko dreifēšanu neitralizē pati dzīve, jo, protams, tam ir skaidri kaitīgs raksturs.
Bibliogrāfiskās atsauces:
- Genetic Drift, khanacademy.org. Saņemts 23. oktobrī plkst https://es.khanacademy.org/science/ap-biology/natural-selection/population-genetics/a/genetic-drift-founder-bottleneck#:~:text=La%20deriva%20g%C3%A9nica%20sucede%20en, 0% 25% 20% 2C% 20de% 20citi% 20alelos.
- Eguiarte, L., Agirra-Plantera, E., Šeinvara, E., Gonsalē, A. un Souza, V. (2010). Gēnu plūsma, populāciju diferenciācija un ģenētiskā struktūra ar piemēriem Meksikas augu sugām. Meksikas Nacionālās autonomās universitātes Ekoloģijas institūta Evolūcijas ekoloģijas departamenta molekulārās un eksperimentālās evolūcijas laboratorija, 1.-30.
- Futuyma, D. Dž. (1992). Evolūcijas bioloģija (sēj. 2). 2. ed. Ribeirão Preto: SBG.