
גלוטמט (נוירוטרנסמיטר): הגדרה ותפקודים
ה גלוטמט מתווך את רוב הסינפסות המעוררות במערכת העצבים המרכזית (CNS). זהו המתווך העיקרי של מידע חושי, מוטורי, קוגניטיבי ורגשי והוא מעורב בהיווצרות זיכרונות ובהחלמתם, בהיותו נוכח ב- 80-90% מהסינפסות המוחיות.
כאילו כל זה מעט זכות, הוא מתערב גם בנוירופלסטיות, בתהליכי למידה והוא מבשרם של גאבא - המוליך העצבי העיקרי המעכב של מערכת העצבים המרכזית. מה עוד תוכלו לבקש ממולקולה?
מה זה גלוטמט?
יִתָכֵן היה אחד מהמעבירים העצביים הנחקרים ביותר במערכת העצבים. בשנים האחרונות המחקר שלו גדל בגלל הקשר שלו עם פתולוגיות נוירו-ניווניות שונות (כגון מחלת אלצהיימר), שהפכה אותו למטרה חזקה לתרופות במחלות שונות.
ראוי גם להזכיר כי בהתחשב במורכבות הקולטנים שלו, זהו אחד הנוירוטרנסמיטרים המסובכים ביותר שנחקרים.
תהליך הסינתזה
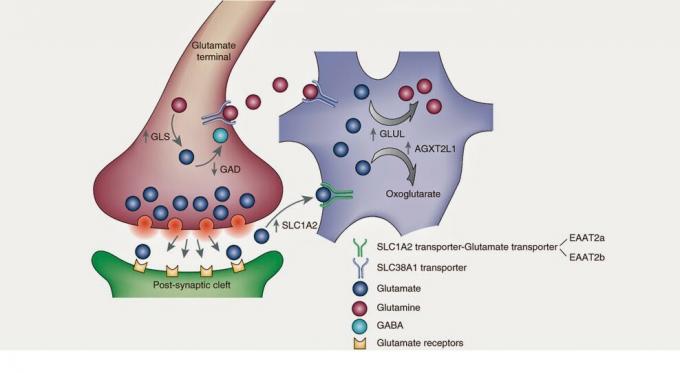
תהליך סינתזת הגלוטמט מתחיל במחזור קרבס, או במחזור החומצה הטריקרבוקסילית. מחזור קרבס הוא מסלול מטבולי או, להבנתנו, רצף של תגובות כימיות במטרה לייצר נשימה תאית במיטוכונדריה. ניתן להבין מחזור מטבולי כמנגנון של שעון, בו כל הילוך ממלא א תפקוד וכשל פשוט של חלק עלול לגרום לפגיעה בשעון או לא שָׁעָה. המחזורים בביוכימיה זהים. מולקולה, באמצעות תגובות אנזימטיות מתמשכות - גלגלי השיניים של השעון - משנה את צורתה והרכבה במטרה להוליד פונקציה תאית. מבשר הגלוטמט העיקרי יהיה אלפא-קטוגלוטרט, אשר יקבל קבוצת אמינו על ידי טרנסמינציה כדי להפוך לגלוטמט.
כדאי להזכיר גם מבשר די משמעותי למדי: גלוטמין. כאשר התא משחרר גלוטמט לחלל החוץ תאי, אסטרוציטים - סוג של תא גליה - התאושש בגלוטמט זה אשר באמצעות אנזים הנקרא גלוטמין סינטטאז יהפוך גלוטמין. יותר מאוחר, גלוטמין משתחרר על ידי אסטרוציטים, שמוחזר על ידי נוירונים כדי להפוך אותם חזרה לגלוטמט. ואולי יותר מאחד ישאל את הדברים הבאים: ואם עליהם להחזיר גלוטמין לגלוטמט בתאי העצב, מדוע האסטרוציט ממיר גלוטמט מסכן לגלוטמין? ובכן, גם אני לא יודע. אולי זה שהאסטרוציטים והנוירונים לא מסכימים או שאולי זה מדעי המוח זה כל כך מסובך בכל מקרה, רציתי לעשות סקירה של אסטרוציטים מכיוון ששיתוף הפעולה שלהם מהווה 40% מה- מַחזוֹר גלוטמט, כלומר רוב הגלוטמט נוצר על ידי תאי גליה אלה.
ישנם מקדימים אחרים ומסלולים אחרים שבאמצעותם ניתן להחזיר גלוטמט המשתחרר לחלל החוץ-תאי. לדוגמא, ישנם נוירונים המכילים טרנספורטר גלוטמט ספציפי –EAAT1 / 2– המחלים ישירות את הגלוטמט לנוירון ומאפשרים לסיים את האות המעורר. להמשך מחקר על סינתזה ומטבוליזם של גלוטמט, אני ממליץ לקרוא את הביבליוגרפיה.
קולטני גלוטמט
כפי שהם מלמדים אותנו בדרך כלל, לכל משדר עצבי יש את הקולטנים שלו בתא הפוסט-סינפטי. הקולטנים, הממוקמים על קרום התא, הם חלבונים אליהם נקשר נוירוטרנסמיטר, הורמון נוירופפטיד וכו ', כדי ליצור סדרה של שינויים בחילוף החומרים הסלולרי של התא בו הוא נמצא הקולטן. בתאי עצב אנו בדרך כלל מציבים את הקולטנים על תאים פוסט-סינפטיים, אם כי זה לא באמת צריך להיות ככה.
הם גם מלמדים אותנו בדרך כלל בשנה הראשונה למרוץ שיש שני סוגים עיקריים של קולטנים: יונוטרופי ומטא-בוטרופי. יונוטרופים הם כאלה שבהם כאשר הליגנד שלהם - "המפתח" של הקולטן - נקשר, הם פותחים תעלות המאפשרות מעבר של יונים לתא. לעומת זאת, מטאוטרופיקה, כאשר הליגנד נקשר, גורמות לשינויים בתא באמצעות שליחים שניים. בסקירה זו אדבר על הסוגים העיקריים של קולטני גלוטמט יונוטרופיים, אם כי אני ממליץ ללמוד בספרות כדי להבין קולטנים מטבוטרופיים. להלן הקולטנים העיקריים היונוטרופיים:
- מקלט NMDA.
- מקלט AMPA.
- לוכד קאינאדו.
קולטני NMDA ו- AMPA והקשר ההדוק שלהם
ההערכה היא ששני סוגי הקולטנים הם מקרומולקולות המורכבות מארבעה תחומים טרנס-ממברניים - כלומר הם מורכבים מארבע יחידות משנה ש הם חוצים את שכבת השומנים השומנית של קרום התא - ושניהם קולטני גלוטמט הפותחים תעלות קטיון - יונים טעונים חיובי. אך למרות זאת, הם שונים משמעותית.
אחד ההבדלים ביניהם הוא הסף שבו הם מופעלים. ראשית, קולטני AMPA הם הרבה יותר מהירים להפעלה; בעוד שלא ניתן להפעיל קולטני NMDA עד שלנוירון יש פוטנציאל קרום של כ -50mV - נוירון כאשר הוא מושבת הוא בדרך כלל סביב -70mV-. שנית, שלב הקטיונים יהיה שונה בכל מקרה. קולטני AMPA ישיגו פוטנציאל ממברנה גבוה בהרבה מקולטני NMDA, שישתפו פעולה בצורה צנועה הרבה יותר. בתמורה, קולטני NMDA ישיגו הפעלות מתמשכות הרבה יותר לאורך זמן מאשר קולטני AMPA. לָכֵן, אלה של AMPA מופעלים במהירות ומייצרים פוטנציאלים מעוררים חזקים יותר, אך מבוטלים במהירות. ולאלה של NMDA לוקח זמן להפעיל, אבל הם מצליחים לשמור על הפוטנציאלים המסעירים שהם מייצרים הרבה יותר זמן.
כדי להבין את זה טוב יותר, בואו נדמיין שאנחנו חיילים ושהנשק שלנו מייצג את הקולטנים השונים. בואו נדמיין שהמרחב החוץ-תאי הוא תעלה. יש לנו שני סוגים של כלי נשק: אקדח ורימונים. רימונים פשוטים ומהירים לשימוש: אתה מסיר את הטבעת, זורק אותה ומחכה להתפוצצותה. יש להם פוטנציאל הרסני רב, אבל ברגע שזרקנו את כולם זה נגמר. האקדח הוא כלי נשק שלוקח לו זמן להעמיס כי צריך להסיר את התוף ולשים את הכדורים בזה אחר זה. אך לאחר שהעמסנו אותו יש לנו שש יריות איתן אנו יכולים לשרוד לזמן מה, אם כי עם פוטנציאל הרבה פחות מרימון. אקדחי המוח שלנו הם קולטני NMDA והרימונים שלנו הם קולטני AMPA.
גלוטמט עודפים וסכנותיהם
הם אומרים כי עודף שום דבר לא טוב ובמקרה של גלוטמט זה נכון. לאחר מכן נביא כמה פתולוגיות ובעיות נוירולוגיות בהן קשור עודף גלוטמט.
1. אנלוגים של גלוטמט עלולים לגרום לרעילות אקסוטו
תרופות אנלוגיות לגלוטמט - כלומר הן ממלאות את אותה פונקציה כמו גלוטמט - כגון NMDA - שקולטן NMDA מקבל את שמו - יכול לגרום לתופעות ניווניות ניווניות במינונים גבוהים באזורי המוח הפגיעים ביותר כגון הגרעין הקשת של ההיפותלמוס. המנגנונים המעורבים בניוון עצבי זה הם מגוונים וכוללים סוגים שונים של קולטני גלוטמט.
2. כמה נוירוטוקסינים שאנחנו יכולים לבלוע בתזונה מפעילים מוות עצבי באמצעות עודף גלוטמט
רעלים שונים של כמה בעלי חיים וצמחים מפעילים את השפעותיהם במסלולי העצב הגלוטמט. דוגמה לכך היא הרעל מזרעי Cycas Circinalis, צמח רעיל שאנו יכולים למצוא באי האוקיאנוס השקט גואם. רעל זה גרם לשכיחות גבוהה של טרשת לרוחב אמיוטרופית באי הזה בו תושביו בלעו אותו מדי יום, מתוך אמונה שהוא שפיר.
3. גלוטמט תורם למוות עצבי איסכמי
גלוטמט הוא הנוירוטרנסמיטר העיקרי בהפרעות מוחיות חריפות כגון התקף לב, דום לב, היפוקסיה לפני הלידה. באירועים אלה בהם חסר חמצן ברקמת המוח, הנוירונים נותרים במצב של דפולריזציה קבועה; בשל תהליכים ביוכימיים שונים. זה מוביל לשחרור קבוע של גלוטמט מהתאים, עם הפעלה מתמשכת של קולטני גלוטמט לאחר מכן. קולטן ה- NMDA חדיר במיוחד לסידן בהשוואה לקולטנים יונוטרופיים אחרים, עודף סידן מוביל למוות עצבי. לכן, היפראקטיביות של קולטנים גלוטמטרגיים מובילה למוות עצבי בגלל סידן תוך-עצבי מוגבר.
4. אֶפִּילֶפּסִיָה
הקשר בין גלוטמט לאפילפסיה מתועד היטב. פעילות אפילפטית נחשבת כקשורה במיוחד לקולטני AMPA, אם כי ככל שמתקדמת לאפילפסיה, קולטני NMDA הופכים חשובים.
האם גלוטמט טוב? האם גלוטמט רע?
בדרך כלל, כאשר קוראים סוג זה של טקסט, בסופו של דבר הוא מאניש את המולקולות בכך שהוא שם עליהן תוויות כ"טובות "או" רעות "- שם זה נקרא אנתרופומורפיזם, חזרה מאוד אופנתית בימי הביניים. המציאות רחוקה למדי מפסקי הדין הפשטניים הללו.
בחברה בה יצרנו מושג של "בריאות" קל לחלק ממנגנוני הטבע להטריד אותנו. הבעיה היא שהטבע לא מבין "בריאות". יצרנו זאת באמצעות רפואה, תעשיות תרופות ופסיכולוגיה. זהו מושג חברתי, וכמו כל המושגים החברתיים הוא נתון לקידום חברות, בין אם זה אנושי או מדעי. ההתקדמות מראה כי גלוטמט קשור למספר פתולוגיות כמו אלצהיימר או סכִיזוֹפרֶנִיָה. זו אינה עין רעה של אבולוציה לאדם, אלא מדובר בחוסר התאמה ביוכימי של מושג שהטבע עדיין לא מבין: החברה האנושית במאה ה -21.
וכמו תמיד, מדוע ללמוד זאת? במקרה זה אני חושב שהתשובה ברורה מאוד. בשל התפקיד שיש לגלוטמט בפתולוגיות ניווניות שונות, הוא גורם למטרה תרופתית חשובה - אם כי גם מורכבת.. כמה דוגמאות למחלות אלה, אם כי לא דיברנו עליהן בסקירה זו מכיוון שאני חושב שניתן היה לכתוב על כך ערך בלעדי, הם מחלת אלצהיימר ו סכִיזוֹפרֶנִיָה. באופן סובייקטיבי, אני מוצא את החיפוש אחר תרופות חדשות עבור סכיזופרניה משתי סיבות עיקריות: שכיחות המחלה ועלות הבריאות נושא; וההשפעות השליליות של תרופות אנטי-פסיכוטיות עכשוויות, אשר במקרים רבים מפריעות לדבקות בטיפול.
טקסט תוקן ונערך על ידי פרדריק מונינט פיי
הפניות ביבליוגרפיות:
ספרים:
- סיגל, ג ' (2006). נוירוכימיה בסיסית. אמסטרדם: אלסבייה.
מאמרים:
- סיטרי, א. ומלנקה, ר. (2007). פלסטיות סינפטית: צורות מרובות, פונקציות ומנגנונים. נוירופסיכופרמקולוגיה, 33 (1), 18-41. http://dx.doi.org/10.1038/sj.npp.1301559
- הרדינגהם, ג '. & Bading, H. (2010). איתות קולטן NMDA חוץ סינפטי: השלכות על הפרעות ניווניות. סקירות טבע מדעי המוח, 11 (10), 682-696. http://dx.doi.org/10.1038/nrn2911
- הרדינגהם, ג '. & Bading, H. (2010). איתות קולטן NMDA חוץ סינפטי: השלכות על הפרעות ניווניות. סקירות טבע מדעי המוח, 11 (10), 682-696. http://dx.doi.org/10.1038/nrn2911
- Kerchner, G. וניקול, ר. (2008). סינפסות שקטות והופעתו של מנגנון פוסט-סינפטי ל- LTP. סקירות טבע מדעי המוח, 9 (11), 813-825. http://dx.doi.org/10.1038/nrn2501
- פפוין, ט. ואולייט, ש. (2014). ארגון, בקרה ותפקוד של קולטני NMDA חוץ סינפטיים. עסקאות פילוסופיות של החברה המלכותית ב ': מדעים ביולוגיים, 369 (1654), 20130601-20130601. http://dx.doi.org/10.1098/rstb.2013.0601