
Генетски помак: шта је то и како утиче на биолошку еволуцију?
Биолошка еволуција, замишљена као скуп промена наследних карактеристика у живих бића кроз генерације, покретач је самог живота и прилагођавања новом окружења.
Варијације унутар популације живих бића настале су услед низа мутација у ДНК које се дешавају насумично, односно не реагује на свесни механизам. Упркос томе, поступци селекције за ове мутације могу бити случајни, или напротив, имају потпуно утемељено објашњење.
Дакле, еволуција је сила која се одржава случајним мутацијама, генетском рекомбинацијом током сексуална репродукција и проток гена (улазак нових чланова у популацију), поред многих других фактора. Један од ових фактора од суштинске важности који често измиче општем разумевању је термин који нас овде занима: генетско одступање. Овде објашњавамо све о овом фасцинантном процесу.
- Повезани чланак: „Теорија биолошке еволуције“
Шта је генетски дрифт?
Прво, овај сложени појам можемо дефинисати као „еволуциону силу која делује заједно са природном селекцијом променом фреквенција алела врста током времена“. Као предговор треба напоменути да
то је стохастички процес, односно догађа се услед случајних или спорадичних неусклађених ефеката.Поред ове почетне карактеристике, још један од појмова који дефинишу генетски помак је и губитак, будући да је ово селекциона сила промовише промене алела у популацији, поправљајући неке и промовишући нестанак други. Детаљније ћемо то видети у следећим редовима.
О алелима и популацији
Поједностављујући генетске појмове, можемо потврдити да је алел свака од варијација које исти ген може представити. Класичан пример за то је семе грашка које је Мендел користио да објасни генетску сегрегацију међу генерацијама. Доминантан „А“ алел може да кодира зелену боју семена, док рецесивни „А“ алел кодира жуту боју.
Како је већина животиња диплоидна бића (која у свом језгру имају два комплета хомологних хромозома), сваки од два кодирајућа алела лика долази од оца односно мајке, због чега би у овом случају могуће варијације биле следеће: АА, Аа и аа. Дакле, ако схватимо да појединац наслеђује два алела за сваки ген, доћи ће до његовог фенотипа (спољне карактеристике) директно кодирани њиховим генотипом (алелне комбинације у њиховом геному), који се наслеђује као комбинација Његови родитељи.
Друго, појам „становништво“ треба мало истражити у сфери биологије, будући да генетски нанос делује на популације, а не на саму врсту. Врста је „затворени“ концепт, јер не може да размењује гене са другим различитим ентитетима. С друге стране, популација је замишљена као „отворени“ одељак, јер различити чланови других чланова могу ући. популације, али исте врсте и размножавају се међу њима, догађај који ће бити од виталног значаја у линијама касније. Једном када успоставимо оба појма на општи начин, спремни смо да схватимо основу генетског заноса.
- Можда ће вас занимати: „Спецификација: шта је то и како се развија у биолошкој еволуцији“
Теоријске основе дрифта
Време је да се држите седишта, јер облине и термини постају мало сложени за објашњење. Генетско одступање одређује се варијансом алелне фреквенције, односно променљивошћу карактера у односу на средњу вредност. Тако ову еволуциону силу можемо израчунати помоћу следеће формуле:
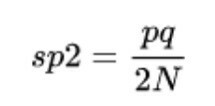
- сп2 одговара варијанси фреквенција алела популација, односно самом генетском заносу.
- п и к су фреквенције алела две популације за лик.
- Н је број јединки унутар сваке од две популације.
Наравно, сваки од ових параметара добија се путем сложених формула, тако да се нећемо више фокусирати на математички темељ ове еволуционе силе. Ако идеја мора бити јасна након читања ових редова, то је следеће: што је мања величина популације, то ће већи генетски помак имати над својим члановима.
Ефективна величина популације
У претходни пасус увели смо кључни термин: величина становништва. Истина је да, узимајући у обзир величину генетског заноса, научници не вреде само да броје појединце у популацији. У овим случајевима број животиња које се у њему репродукују мора бити поуздано квантификован.
Врло јасан пример разлике између укупне популације и ефективне величине популације су демографске студије водоземаца. На пример, уобичајену популацију крастача може чинити 120 чланова. Ако прибегнемо генетској анализи, можемо приметити да се, сигурно, годишње репродукује само око 40 одраслих одраслих особа, остављајући максимално потомство. Дакле, ефективна величина популације (Не) која би претрпела ефекте заношења била би 40, а не 120.
Ефекти генетског заноса
Генетски помак има неколико ефеката на популације живих бића, али их можемо поделити у два велика блока:
- Производи промену фреквенција алела у популацији. То може значити да се они повећавају или смањују, јер је то ствар чисте шансе.
- Смањује дугорочне генетске варијације у популацијама.
Ова последња тачка је од суштинске важности, јер генетски занос смањује варијабилност, што се на крају преводи у већу рањивост становништва на промене у окружењу. Узмимо практичан пример.
Ако имамо у фиктивној популацији од 10 птица, 8 црвених и 2 жуте, природно је мислити да је, сасвим случајно, вероватније него у следеће генерације, црвени чланови изгледају заступљенији (јер ако се од тих 10 репродукују само 3, постоји могућност да су сва 3 обојена Црвена). У првој генерацији, алелна фреквенција црвеног знака „п“ била би 0,8, док би жути знак „к“ имао фреквенцију 0,2.
Ако се у случају репродукују само 3 мушкарца и женке црвене боје, теоретски би алел к могао да нестане следеће генерације, па је п = 1 и к = 0, сви потомци су црвени (знак п би био комплет). Ово је стварни ефекат генетског заноса, који случајно ствара фиксацију најраспрострањенијих ликова у популацији и на крају одбацује најјединствене.
Спас становништва
Срећом, имамо силу која у великој мери избегава ову случајну селекцију: природну селекцију. У овом случају, суочени смо са еволуционим мотором који уопште не одговара случајним и стохастичким процесима, будући да особине сваког појединца могу одредити њихов опстанак, репродукцију и последичну заступљеност у будућим генерацијама.
Такође треба напоменути да је горе наведени пример прилично шепав због самонаметнутог редукционизма, будући да је очигледно да су многи морфолошки знакови кодирани са више од једног гена (као што је боја очију, јер пример). Даље, у популацији од 1000 јединки, а не 10, јасно је да је нестанак алела много сложенији од његовог „брисања“ у једној генерацији.
С друге стране, проток гена је још један кључни концепт који избегава ефекте генетског заноса. Алел би могао да се фиксира у популацији током времена, али ако се појаве нови чланови са различитим алелима и репродукују са јединкама почетне популације, обновљена генетска варијабилност уводи се у следеће генерације.
Коначно, то је неопходно ограничити мутације се јављају случајно у живим бићима. Дакле, у ДНК могу настати варијације које кодирају нове алеле, због чега (на мање теоретски) у затвореној популацији нови знакови се могу и даље појављивати у а спорадично.
Резиме
Као што смо видели, генетски помак то је главни еволутивни мотор живих бића заједно са природном селекцијом, али се разликује од потоњег због своје случајности и случајности. Са чисто теоријске тачке гледишта, ако није било догађаја као што су проток гена, појава мутација или природном селекцијом, све популације би на крају имале по један алел за сваки ген, чак и ако би требало много генерације.
То се, наравно, доводи до мање генетске варијабилности, што значи лошији одговор популације и нивоа појединца на промене и неприкладност околине. Дакле, генетском заносу се супротставља сам живот, јер он, наравно, има јасан штетан карактер.
Библиографске референце:
- Генетички нанос, кханацадеми.орг. Подигнуто 23. октобра у https://es.khanacademy.org/science/ap-biology/natural-selection/population-genetics/a/genetic-drift-founder-bottleneck#:~:text=La%20deriva%20g%C3%A9nica%20sucede%20en, 0% 25% 20% 2Ц% 20де% 20остал% 20алелос.
- Егуиарте, Л., Агуирре-Плантер, Е., Сцхеинвар, Е., Гонзалез, А., & Соуза, В. (2010). Проток гена, диференцијација и генетска структура популација, са примерима у мексичким биљним врстама. Лабораторија за молекуларну и експерименталну еволуцију, Одељење за еволуциону екологију, Институт за екологију, Национални аутономни универзитет у Мексику, 1-30.
- Футуима, Д. Ј. (1992). Еволуциона биологија (књ. 2). 2. изд. Рибеирао Прето: СБГ.