
La dérive génétique: qu'est-ce que c'est et comment affecte-t-elle l'évolution biologique ?
L'évolution biologique, conçue comme l'ensemble des modifications des caractéristiques héréditaires êtres vivants à travers les générations, est le moteur de la vie elle-même et de l'adaptation aux nouvelles environnements.
La variation au sein d'une population d'êtres vivants est donnée par une série de mutations de l'ADN qui se produisent de manière aléatoire, c'est-à-dire qu'elle ne répond pas à un mécanisme conscient. Même ainsi, les processus de sélection de ces mutations peuvent être aléatoires, ou au contraire, avoir une explication pleinement fondée.
Ainsi, l'évolution est une force entretenue par des mutations aléatoires, des recombinaisons génétiques au cours de reproduction sexuée et flux de gènes (entrée de nouveaux membres dans une population), parmi de nombreux autres facteurs. L'un de ces facteurs d'importance essentielle qui échappe souvent à l'entendement général est le terme qui nous intéresse ici: dérive génétique. Ici, nous vous expliquons tout sur ce processus fascinant.
- Article associé: "La théorie de l'évolution biologique"
Qu'est-ce que la dérive génétique ?
Premièrement, nous pouvons définir ce terme complexe comme « une force évolutive qui agit en conjonction avec la sélection naturelle en modifiant les fréquences alléliques des espèces au fil du temps ». En guise de préface, il convient de noter que c'est un processus stochastique, c'est-à-dire qu'il se produit en raison d'effets aléatoires ou sporadiques non corrélés.
En plus de cette caractéristique initiale, un autre des termes qui définissent la dérive génétique est la perte, puisque cette La force de sélection favorise la variation des allèles dans la population, en fixant certains et favorisant la disparition des autres. Nous le verrons plus en détail dans les lignes suivantes.
À propos des allèles et de la population
Simplifier les termes génétiques, on peut affirmer qu'un allèle est chacune des variations que peut présenter un même gène. Un exemple classique est celui des graines de pois utilisées par Mendel pour expliquer la ségrégation génétique à travers les générations. Un allèle dominant "A" peut coder pour une couleur de graine verte, tandis qu'un allèle "a" récessif code pour une couleur jaune.
Comme la plupart des animaux sont des êtres diploïdes (qui ont deux ensembles de chromosomes homologues dans leur noyau), chacun des deux allèles codants d'un caractère proviendra respectivement du père et de la mère, c'est pourquoi les variations possibles dans ce cas seraient les suivantes: AA, Aa et aa. Ainsi, si l'on comprend qu'un individu hérite de deux allèles pour chaque gène, son phénotype (caractéristiques externes) viendra directement codés par leur génotype (combinaisons alléliques dans leur génome), qui est hérité comme une combinaison de ceux de ses parents.
Deuxièmement, le terme «population» doit être exploré un peu dans le domaine de la biologie, car la dérive génétique agit sur les populations et non sur l'espèce elle-même. Une espèce est un concept "fermé", car elle ne peut pas échanger de gènes avec d'autres entités différentes. D'autre part, une population est conçue comme un compartiment « ouvert », puisque différents membres d'autres membres peuvent entrer. populations mais de la même espèce et se reproduisent entre elles, un événement qui sera d'une importance vitale dans les lignées plus tard. Une fois que nous avons établi les deux termes d'une manière générale, nous sommes prêts à comprendre le fondement de la dérive génétique.
- Vous etes peut etre intéressé: "La spéciation: qu'est-ce que c'est et comment elle se développe dans l'évolution biologique"
Fondement théorique de la dérive
Il est temps de s'accrocher au siège, car les courbes et les termes sont un peu complexes à expliquer. Dérive génétique est déterminé par la variance de la fréquence allélique, c'est-à-dire la variabilité des caractères par rapport à la moyenne. Ainsi, nous pouvons calculer cette force évolutive en utilisant la formule suivante :
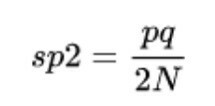
- sp2 correspond à la variance des fréquences alléliques des populations, c'est-à-dire à la dérive génétique elle-même.
- p et q sont les fréquences alléliques de deux populations pour un caractère.
- N est le nombre d'individus au sein de chacune des deux populations.
Bien entendu, chacun de ces paramètres est obtenu grâce à des formules complexes, nous n'allons donc pas nous concentrer davantage sur le fondement mathématique de cette force évolutive. Si une idée doit être claire après avoir lu ces lignes, c'est la suivante: plus la taille de la population est petite, plus la dérive génétique aura de pouvoir sur ses membres.
La taille effective de la population
Nous avons introduit un terme clé dans le paragraphe précédent: la taille de la population. La vérité est que, compte tenu de l'ampleur de la dérive génétique, les scientifiques ne valent pas seulement la peine de compter les individus dans une population. Dans ces cas le nombre d'animaux qui s'y reproduisent doit être quantifié de manière fiable.
Les études démographiques sur les amphibiens sont un exemple très clair de la différence entre la population totale et la taille effective de la population. Une population de crapauds communs, par exemple, peut être composée de 120 membres. Si nous recourons à l'analyse génétique, nous pouvons observer que, sûrement, seulement environ 40 adultes au total se reproduisent chaque année, laissant un maximum de progéniture. Ainsi, la taille effective de la population (Ne) qui subirait les effets de la dérive serait de 40 et non de 120.
Les effets de la dérive génétique
La dérive génétique a plusieurs effets sur les populations d'êtres vivants, mais on peut les diviser en deux grands blocs :
- Produit un changement dans les fréquences alléliques au sein de la population. Cela peut signifier que ceux-ci augmentent ou diminuent, car c'est une question de pur hasard.
- Réduit la variation génétique à long terme dans les populations.
Ce dernier point est d'une importance capitale, car la dérive génétique diminue la variabilité, ce qui se traduit finalement par une plus grande vulnérabilité de la population aux changements environnementaux. Prenons un exemple pratique.
Si nous avons dans une population fictive de 10 oiseaux, 8 rouges et 2 jaunes, il est naturel de penser que, par pur hasard, il est plus probable que dans le génération suivante, les membres rouges semblent plus représentés (car si sur ces 10 seuls 3 sont reproduits, il est possible que tous les 3 soient colorés Rouge). Dans la première génération, la fréquence allélique du caractère rouge "p" serait de 0,8, tandis que le caractère jaune "q" aurait une fréquence de 0,2.
Si seulement 3 mâles et femelles de couleur rouge se reproduisent dans un événement, théoriquement l'allèle q pourrait disparaître dans la génération suivante, donc p = 1 et q = 0, tous les descendants étant rouges (le caractère p aurait été ensemble). C'est l'effet réel de la dérive génétique, qui par hasard, il produit une fixation des caractères les plus distribués dans la population et finit par rejeter les plus uniques.
Le salut des populations
Heureusement, nous avons une force qui évite largement cette sélection aléatoire: la sélection naturelle. Dans ce cas, nous sommes face à un moteur évolutif qui ne correspond pas du tout à des processus aléatoires et stochastiques, puisque les caractéristiques de chaque individu peuvent déterminer sa survie, sa reproduction et sa représentation conséquente dans les générations futures.
Il convient également de noter que l'exemple cité ci-dessus est assez boiteux en raison du réductionnisme auto-imposé, étant donné qu'il est clair que de nombreux caractères morphologiques sont codés par plus d'un gène (comme la couleur des yeux, par exemple Exemple). De plus, dans une population de 1000 individus plutôt que 10, il est clair que la disparition d'un allèle est bien plus complexe que son « effacement » en une seule génération.
D'un autre côté, le flux génétique est un autre concept clé qui évite les effets de la dérive génétique. Un allèle pourrait être fixé dans une population au fil du temps, mais si de nouveaux membres apparaissent avec différents allèles et reproduire avec les individus de la population initiale, une variabilité génétique renouvelée est introduite dans la suite générations.
Enfin, il faut limiter les mutations se produisent au hasard chez les êtres vivants. Ainsi, des variations peuvent survenir dans l'ADN qui code pour de nouveaux allèles, c'est pourquoi (au moins théoriquement) dans une population fermée, de nouveaux caractères peuvent continuer à apparaître dans un sporadique.
résumé
Comme nous l'avons vu, la dérive génétique c'est le principal moteur de l'évolution des êtres vivants avec la sélection naturelle, mais il diffère de ce dernier par son caractère aléatoire et aléatoire. D'un point de vue purement théorique, s'il n'y avait pas d'événements tels que des flux de gènes, l'apparition de mutations ou la sélection naturelle, toutes les populations finiraient par avoir un seul allèle pour chaque gène, même s'il fallait plusieurs générations.
Ceci, naturellement, se traduit par moins de variabilité génétique, ce qui signifie une plus mauvaise réponse au niveau de la population et de l'individu aux changements et aux intempéries de l'environnement. Ainsi, la dérive génétique est contrecarrée par la vie elle-même, car bien sûr, elle a un caractère délétère évident.
Références bibliographiques:
- Dérive génétique, khanacademy.org. Reçu le 23 octobre à https://es.khanacademy.org/science/ap-biology/natural-selection/population-genetics/a/genetic-drift-founder-bottleneck#:~:text=La%20deriva%20g%C3%A9nica%20sucede%20en, 0% 25% 20% 2C% 20de% 20autre% 20alelos.
- Eguiarte, L., Aguirre-Planter, E., Scheinvar, E., González, A., & Souza, V. (2010). Flux de gènes, différenciation et structure génétique des populations, avec des exemples d'espèces végétales mexicaines. Laboratoire d'évolution moléculaire et expérimentale, Département d'écologie évolutive, Institut d'écologie, Université nationale autonome du Mexique, 1-30.
- Futuyma, D. J. (1992). Biologie évolutive (Vol. 2). 2. éd. Ribeirão Preto: SBG.
