
Glutamat (nevrotransmitter): definisjon og funksjoner
De glutamat formidler de fleste eksitatoriske synapsene i sentralnervesystemet (CNS). Det er hovedformidleren for sensorisk, motorisk, kognitiv og emosjonell informasjon og er involvert i dannelsen av minner og deres gjenoppretting, og er til stede i 80-90% av hjernesynapsene.
Som om alt dette er av liten fortjeneste, griper det også inn i nevroplastisitet, læringsprosesser og er forløperen til GABA –Hovedhemmende nevrotransmitter av CNS. Hva mer kan du be om et molekyl?
Hva er glutamat?
Muligens har vært en av de mest studerte nevrotransmitterne i nervesystemet. De siste årene har studien økt, på grunn av forholdet til forskjellige nevrodegenerative patologier (for eksempel Alzheimers sykdom), som har gjort det til et kraftig legemiddelmål i forskjellige sykdommer.
Det er også verdt å nevne at gitt reseptorers kompleksitet, er dette en av de mest kompliserte nevrotransmitterne å studere.
Synteseprosessen
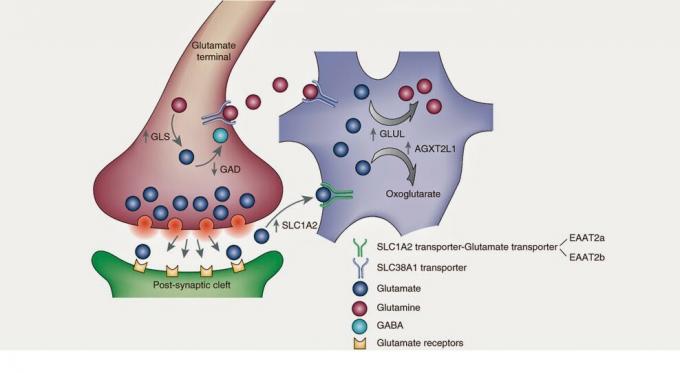
Glutamatsyntese-prosessen begynner i Krebs-syklusen, eller trikarboksylsyresyklusen. Krebs-syklusen er en metabolsk vei eller, for oss å forstå,
en rekke kjemiske reaksjoner for å produsere cellulær respirasjon i mitokondriene. En metabolsk syklus kan forstås som mekanismen til en klokke, der hvert gir oppfyller a funksjon og enkel svikt i en del kan føre til at klokken blir skadet eller ikke time. Syklusene i biokjemi er de samme. Et molekyl endrer form og sammensetning gjennom kontinuerlige enzymatiske reaksjoner - klokkens tannhjul - for å gi en cellulær funksjon. Den viktigste glutamatforløperen vil være alfa-ketoglutarat, som vil motta en aminogruppe ved transaminering for å bli glutamat.Det er også verdt å nevne en annen ganske betydelig forløper: glutamin. Når cellen frigjør glutamat i det ekstracellulære rommet, astrocytter - en type celle glial - gjenopprett dette glutamatet som, gjennom et enzym som kalles glutaminsyntetase, blir glutamin. Seinere, glutamin frigjøres av astrocytter, som gjenvinnes av nevroner for å bli transformert tilbake til glutamat. Og muligens vil mer enn en spørre følgende: Og hvis de må returnere glutamin til glutamat i nevronen, hvorfor omdanner astrocyten dårlig glutamat til glutamin? Jeg vet ikke det heller. Kanskje er det at astrocytter og nevroner ikke er enige, eller kanskje det er at Nevrovitenskap Det er så komplisert I alle fall ønsket jeg å gjøre en gjennomgang av astrocytter fordi deres samarbeid utgjør 40% av omsetning glutamat, som betyr det mesteparten av glutamatet utvinnes av disse gliacellene.
Det er andre forløpere og andre veier der glutamat som frigjøres i det ekstracellulære rommet gjenvinnes. For eksempel er det nevroner som inneholder en spesifikk glutamattransportør –EAAT1 / 2– som direkte gjenoppretter glutamat til nevronet og lar eksitatorisk signal avsluttes. For videre studier av syntesen og metabolismen av glutamat, anbefaler jeg å lese bibliografien.
Glutamatreseptorer
Som de vanligvis lærer oss, hver nevrotransmitter har sine reseptorer på den postsynaptiske cellen. Reseptorene, som ligger på cellemembranen, er proteiner som en nevrotransmitter, hormon, nevropeptid, etc., for å gi opphav til en rekke endringer i cellemetabolismen til cellen der den ligger i reseptoren. I nevroner plasserer vi vanligvis reseptorene på postsynaptiske celler, selv om det egentlig ikke trenger å være slik.
De lærer oss også vanligvis det første året av løpet at det er to hovedtyper av reseptorer: ionotropisk og metabotropisk. Ionotropika er de som når deres ligand - "nøkkelen" til reseptoren - binder, åpner de kanaler som tillater passering av ioner i cellen. Metabotropics derimot, når liganden binder, forårsaker endringer i cellen gjennom andre budbringere. I denne gjennomgangen vil jeg snakke om hovedtyper av ionotrope glutamatreseptorer, selv om jeg anbefaler å studere litteraturen for å forstå metabotrope reseptorer. Her er de viktigste ionotrope reseptorene:
- NMDA-mottaker.
- AMPA-mottaker.
- Kainado-fangst.
NMDA- og AMPA-reseptorer og deres nære forhold
Begge typer reseptorer antas å være makromolekyler som består av fire transmembrane domener - det vil si at de består av fire underenheter som de krysser lipiddobbeltlaget i cellemembranen - og begge er glutamatreseptorer som vil åpne kationkanaler - positivt ladede ioner. Men allikevel er de vesentlig forskjellige.
En av forskjellene deres er terskelen de aktiveres på. For det første er AMPA-reseptorer mye raskere å aktivere; mens NMDA-reseptorer ikke kan aktiveres før nevronet har et membranpotensial på ca. -50mV - et nevron når det inaktiveres er vanligvis rundt -70mV-. For det andre vil kationetrinnet være forskjellig i hvert tilfelle. AMPA-reseptorer vil oppnå mye høyere membranpotensialer enn NMDA-reseptorer, som vil samarbeide mye mer beskjedent. Til gjengjeld vil NMDA-reseptorer oppnå mye mer vedvarende aktiveringer over tid enn AMPA-reseptorer. Derfor, de av AMPA aktiveres raskt og produserer sterkere eksitatoriske potensialer, men deaktiveres raskt. Og de fra NMDA tar tid å aktivere, men de klarer å opprettholde de spennende potensialene de genererer mye lenger.
For å forstå det bedre, la oss forestille oss at vi er soldater og at våpnene våre representerer de forskjellige reseptorene. La oss forestille oss at det ekstracellulære rommet er en grøft. Vi har to typer våpen: revolver og granater. Granater er enkle og raske å bruke: du fjerner ringen, kaster den og venter på at den skal eksplodere. De har mye destruktivt potensiale, men når vi først har kastet dem alle, er det over. Revolveren er et våpen som det tar tid å laste fordi du må fjerne trommelen og sette kulene en etter en. Men når vi først har lastet den, har vi seks skudd som vi kan overleve en stund med, men med mye mindre potensial enn en granat. Hjernens revolvere er NMDA-reseptorer og granatene våre er AMPA-reseptorer.
Glutamat overdreven og deres farer
De sier at i overkant er ingenting bra, og i tilfelle glutamat oppfylles det. Deretter vi vil sitere noen patologier og nevrologiske problemer der et overskudd av glutamat er relatert.
1. Glutamatanaloger kan forårsake eksotoksisitet
Legemidler som er analoge med glutamat - det vil si at de har samme funksjon som glutamat - som NMDA - som NMDA-reseptoren får navnet sitt til - kan forårsake nevrodegenerative effekter ved høye doser i de mest sårbare hjerneområdene slik som den buede kjernen til hypothalamus. Mekanismene som er involvert i denne nevrodegenerasjonen er forskjellige og involverer forskjellige typer glutamatreseptorer.
2. Noen nevrotoksiner som vi kan innta i kostholdet vårt, utøver nevronedød gjennom overflødig glutamat
Ulike giftstoffer fra noen dyr og planter utøver effekten gjennom nervebanene i glutamatet. Et eksempel er giften fra frøene til Cycas Circinalis, en giftig plante som vi kan finne på Stillehavsøya Guam. Denne giften forårsaket en høy forekomst av Amyotrofisk lateral sklerose på denne øya hvor innbyggerne inntok den daglig, og trodde den var godartet.
3. Glutamat bidrar til iskemisk neuronal død
Glutamat er den viktigste nevrotransmitteren ved akutte hjernesykdommer som hjerteinfarkt, hjertestans, pre / perinatal hypoksi. I disse hendelsene der det er mangel på oksygen i hjernevevet, forblir nevronene i en tilstand av permanent depolarisering; på grunn av forskjellige biokjemiske prosesser. Dette fører til permanent frigjøring av glutamat fra cellene, med påfølgende vedvarende aktivering av glutamatreseptorer. NMDA-reseptoren er spesielt permeabel for kalsium sammenlignet med andre ionotrope reseptorer, og overflødig kalsium fører til neuronal død. Derfor fører hyperaktivitet til glutamaterge reseptorer til nevronal død på grunn av økt intraneuronal kalsium.
4. Epilepsi
Forholdet mellom glutamat og epilepsi er godt dokumentert. Epileptisk aktivitet anses å være spesielt relatert til AMPA-reseptorer, men etter hvert som epilepsi utvikler seg, blir NMDA-reseptorer viktige.
Er glutamat bra? Er glutamat dårlig?
Når man leser denne typen tekst, ender han opp med å humanisere molekylene ved å sette etiketter på dem som "gode" eller "dårlige" - som har et navn og kalles antropomorfisme, veldig fasjonabelt tilbake i middelalderen. Virkeligheten er ganske langt fra disse forenklede dommene.
I et samfunn der vi har generert et begrep om "helse", er det lett for noen av naturens mekanismer å plage oss. Problemet er at naturen ikke forstår "helse". Vi har skapt det gjennom medisin, farmasøytisk industri og psykologi. Det er et sosialt begrep, og som alle sosiale begreper er det underlagt samfunnets fremgang, det være seg menneskelig eller vitenskapelig. Fremskritt viser at glutamat er assosiert med en rekke patologier som Alzheimers eller Schizofreni. Dette er ikke et ondt øye med evolusjonen for mennesket, det er snarere et biokjemisk misforhold mellom et konsept som naturen fremdeles ikke forstår: det menneskelige samfunn i det 21. århundre.
Og som alltid, hvorfor studere dette? I dette tilfellet tror jeg svaret er veldig klart. På grunn av rollen som glutamat har i forskjellige nevrodegenerative patologier, resulterer det i et viktig - men også komplekst - farmakologisk mål. Noen eksempler på disse sykdommene, selv om vi ikke har snakket om dem i denne anmeldelsen fordi jeg vurderer det at en oppføring kunne skrives utelukkende om dette, er Alzheimers sykdom og Schizofreni. Subjektivt finner jeg jakten på nye medisiner for schizofreni av to hovedårsaker: utbredelsen av denne sykdommen og helsekostnaden bærer; og bivirkningene av dagens antipsykotika, som i mange tilfeller hindrer overholdelse av terapi.
Tekst korrigert og redigert av Frederic Muniente Peix
Bibliografiske referanser:
Bøker:
- Siegel, G. (2006). Grunnleggende nevrokjemi. Amsterdam: Elsevier.
Artikler:
- Citri, A. & Malenka, R. (2007). Synaptisk plastisitet: Flere former, funksjoner og mekanismer. Nevropsykofarmakologi, 33 (1), 18-41. http://dx.doi.org/10.1038/sj.npp.1301559
- Hardingham, G. & Bading, H. (2010). Synaptisk versus ekstrasynaptisk NMDA-reseptorsignalering: implikasjoner for nevrodegenerative lidelser. Nature Reviews Neuroscience, 11 (10), 682-696. http://dx.doi.org/10.1038/nrn2911
- Hardingham, G. & Bading, H. (2010). Synaptisk versus ekstrasynaptisk NMDA-reseptorsignalering: implikasjoner for nevrodegenerative lidelser. Nature Reviews Neuroscience, 11 (10), 682-696. http://dx.doi.org/10.1038/nrn2911
- Kerchner, G. & Nicoll, R. (2008). Stille synapser og fremveksten av en postsynaptisk mekanisme for LTP. Nature Reviews Neuroscience, 9 (11), 813-825. http://dx.doi.org/10.1038/nrn2501
- Papouin, T. & Oliet, S. (2014). Organisering, kontroll og funksjon av ekstrasynaptiske NMDA-reseptorer. Philosophical Transactions Of The Royal Society B: Biological Sciences, 369 (1654), 20130601-20130601. http://dx.doi.org/10.1098/rstb.2013.0601
